Рассеиваемый при гидролизе полимера олигомерный продукт может быть использован "микрофлорой рассеяния", теперь получившей название диссипотрофов. Эти организмы легче всего выделяются в культуру и лучше всего известны в лабораторной практике. Диссипотрофы представлены в основном одиночными клетками, часто подвижными. В анаэробном сообществе они нагляднее всего представлены спирохетами с очень выгодным отношением поверхности к объему. Зависимость диссипотрофов от гидролитиков очевидна. Менее ясна зависимость гидролитиков от диссипотрофов, связаная с тем, что диссипотрофы снижают концентрацию продуктов гидролиза ниже порогового уровня, останавливающего синтез гидролаз, находящийся под метаболическим контролем ретроингибирования. Необходимость взаимодействия двух групп организмов при разложении даже такого простого вещества, как линейный полимер глюкозы - целлюлоза - лучше рассмотреть на конкретном примере анаэробных термофильных целлюлозолитических организмов.
Разложение целлюлозы представляет важнейший маршрут в наземных и многих морских экосистемах, поскольку целлюлоза образуется в больших количествах как структурный компонент клеточной оболочки растениями и водорослями. Способностью к разложению целлюлозы обладают многие грибы, из которых наиболее подробно была изучена Trichoderma, некоторые актиномицеты, часть которых специализирована на разложении хитина, в том числе грибной стенки, аэробные бактерии, как, например, цитофаги, некоторые миксобактерии, бациллы, целлуломонас, целлвибрио. Среди анаэробов наиболее легкими для исследования объектами оказались клостридии. Знания относительно целлюлозолитических организмов были сильно расширены за счет описания анаэробных неспоровых организмов.
Использование целлюлозы обусловлено взаимодействием с нерастворимым гидрофильным субстратом, представляющим структурно-организованные волокна. Отсюда следуют необходимость физического контакта клеток с нерастворимым субстратом и обрастание его биопленкой. Деградация целлюлозы требует действия внеклеточных ферментов целлюлаз. Доступность для
202
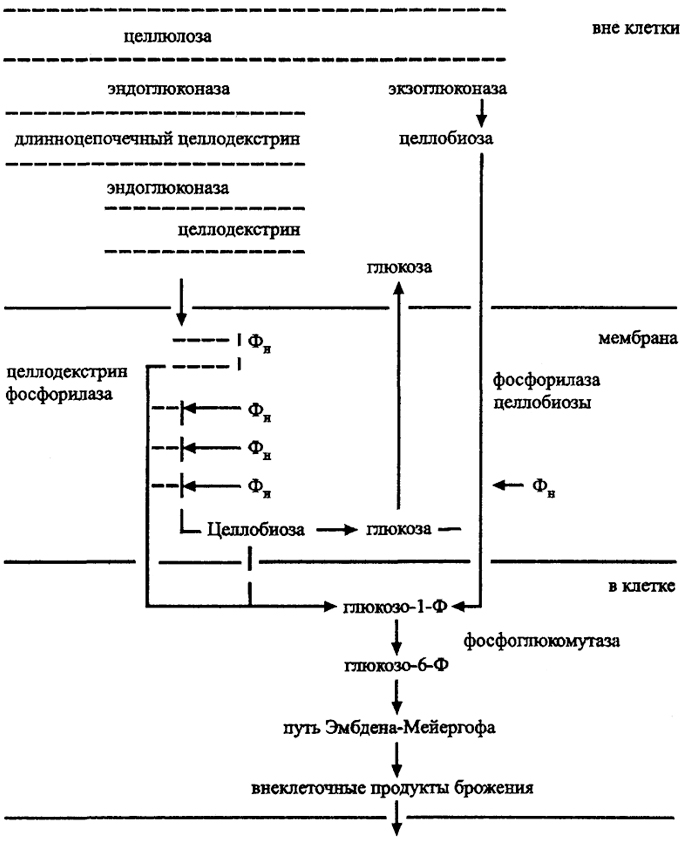
Рис. 33. Использование целлюлозы Clostridium thermocellum
фермента сильно коррелирует с пористостью целлюлозного материала. Физический предел пор для действия фермента составляет величина в 40-50 ангстрем.
Разложение целлюлозы было подробно исследовано на примере Clostridium thermocellum в связи с организацией производства этанола на целлюлозе (рис. 33). Гидролиз осуществляется
203
внеклеточным целлюлазным комплексом (целлюлосомой), включающим ряд ферментов, прежде всего эндоглюконазу, гидролизующую (3-1,4 связь в разных местах с образованием случайных по длине целлодекстринов, которые могут быть далее гидролизова-ны до более коротких олигомеров и поступать в клетку. У С. thermocellum эндоглюконазный комплекс наиболее активен. Менее активна у этого организма экзоглюконаза, отщепляющая с редуцирующего конца полимерной цепочки дисахарид целлобиозу. Соотношение эндоглкжоназа/экзоглюконаза различно у разных организмов. Например, у гриба Trichoderma преобладает экзоглюконаза. Целлюлазный комплекс по-разному ингибируется конечными продуктами у разных целлюлозолитиков.
Продукты гидролиза целлюлозы переносятся в клетку двумя разными путями с участием необычных ферментов: фосфорилазы целлобиозы или целлодекстрин-фосфорилазы. Обе эти реакции фосфорилирования термодинамически обусловлены гидролитическим выбросом глюкозы вне клетки при АТФ-независимом транспорте остающейся части полиглюкозы внутрь клетки. Там она подвергается сбраживанию, по обычному пути брожения, с образованием летучих жирных кислот и, прежде всего, бутирата, ацетата, этанола, Н2. Глюкоза не обязательно используется всеми целлюлозолитиками.
Сходные пути гидролиза действуют и с другими полисахаридами, требуя специфических ферментных систем. Легче всего гидролизуются растворимые полисахариды, для которых есть соответствующие ферменты: крахмал с амилазой, пуллулан с пуллуланазой, ксилан с ксиланазой и т.д.
Из описания действия целлюлазного комплекса с очевидностью следует, что целлюлозолитические организмы должны развиваться в тесных симбиотических отношениях с другими организмами. Прежде всего это относится к необходимости удаления глюкозы диссипотрофами. В исследованной бинарной культуре с С. thermocellum глюкозу использовал Thermoanaerobacter, но можно привести много подобных примеров, таких как пару гало-филов Halocella - Haloanaerobium. Аэробным целлюлозолити-кам цитофагам сопутствуют типичные диссипотрофы просте-кобактерии, как показала Л. В. Васильева, развивающиеся на
204
фильтровальной бумаге в зоне вслед за движущимся фронтом цитофаг. Таким образом, образование пары "гидролитик - диссипотроф" представляет правило. Для анаэробных целлюлозолити-ков необходимо удаление продуктов брожения, так же как для многих первичных анаэробов. Поэтому целлюлозолитические организмы развиваются в тесных синтрофных отношениях с рядом сопутствующих форм.
Помимо катаболических взаимодействий, спутники обеспечивают анаболические нужды целлюлозолитиков. А.А. Имшенецкий обратил особое внимание на симбиотические отношения с азотфиксаторами, снабжавшими целлюлозолитиков связанным азотом, а также различными факторами роста. В результате целлюлозолитическое сообщество взаимодополняющих организмов представляет функционально нечто совершенно иное, чем чистая культура, где необходимо специальными приемами восполнять недостающие потребности организма. Поэтому в каждых экологических условиях формируется своя группировка, связанная с разложением целлюлозы, включающая функционально сходные, но различные по происхождению организмы.
Молекулярные методы позволили разобраться в происхождении целлюлозолитической активности на примере узкой группы экстремально термофильных целлюлозолитиков. Из них наиболее изучен Caldicellulosiruptor, первоначально выделенный из гидротерм Курильских островов под названием "Anaerocellum". Кроме того, есть филогенетически далекие Rhodothermus marinus, Caldibacillus. Гены, определяющие целлюлозолитическую активность, часто полифункциональны, несколько различаются по последовательностям, но вместе с тем существует высокая вероятность того, что латеральный перенос генов имел важное значение в эволюции этой функциональной группы организмов.
205
